多毛类见于各海洋生境,其形态结构的变化是与其栖息地、摄食、生殖等习性密切相关的。
1.生活方式和运动:游走多毛类,常是自由活动的穴居、浮游、爬行者或在生活史一定阶段的管居者;而定居多毛类,成体多为永久性的穴居或管栖者,很少离开栖管或出穴外。
(1)爬行或底表栖居者(crawling polychaete or surface dweller)(图18-1 A~C):常藏于裂隙、石块、贝壳下或海藻、水螅、苔藓及其他固着生物丛中,包括沙蚕、裂虫、叶须虫和鳞沙蚕等。其虫体的典型结构为:头部发达具感觉器官或附肢,疣足大且常用以爬行(图18-9),纵肌强壮。

图18-1 海洋各生境中的多毛动物
A.沙蚕;B.裂虫;C.鳞沙蚕;D.巢沙蚕;E.浮蚕;F.磷虫;G.沙蠋(示生活史);H.笔帽虫;I.蛰龙介;J.缨鳃虫;K.石灰虫;L.螺旋虫

图18-9 沙蚕爬行(A)和游泳(B)(仿Clark)
(2)浮游多毛类(pelagic or planktonic polychaete)(图18-1 E):在水层中浮
游生活,包括眼蚕、浮蚕、盲蚕等。其共同特征为:体透明,头部明显,疣足多叶
片状,纵肌发达,肉食性。
(3)穴居多毛类(burrowing polychaete):适于在泥沙底穴居者,依穴居方式
又分为两类:①地下甬道栖居者(gallery dweller),见于吻沙蚕(图18-12 A)、索
沙蚕(18-12 B)、海蛹(图18-10)和小头虫(图18-13 D)等,可挖掘大面积的穴道
或地下甬道,并有许多孔开于地表,甬道壁为黏液衬里以润滑并防塌陷,其穴居
的适应很似寡毛类的蚯蚓,虫体通常细长,头部尖圆常无附肢或感官,疣足多退
化,环肌发达,隔膜较完整,皆为钻穴所必须,肉食性或直接摄食沉积物。②定
居穴居者(sedentary burrower),见于沙蠋(图18-1 G,图18-13 E)、蛰龙介(图
18-1 I)和丝鳃虫(图18-13 B)等,可在泥沙中建造临时的两端开孔的U形管或
一端开孔的直管,体膨胀呈圆柱状且分区,疣足退化、部分变为具齿片或足刺刚
毛的横脊、这有助于抓住穴壁,头部无附肢或附肢为特化的摄食结构。沙蠋为
直接摄食沉积物者,而蛰龙介则为间接摄食沉积物者。

图18-10 阿曼吉虫(A)的波状运动(B)(仿clark)

图18-11 原环虫(A)以1/2波长运动(B)(十:示波沿体向后传递)(仿clark)
(4)管栖多毛类(tubicolous polychaete):永久生活于虫体分泌的或由外来物
建成的栖管中。在形态和结构上变化很大,具多种适应类群。①游走管栖类(errantic tubicolou):如矶沙蚕(图18-12 F)和巢沙蚕(图18-1 D),头部感官和附肢发
达,疣足不明显退化,栖管直立,由沙粒和有机物质粘成。潮间带泥沙滩习见的巢
沙蚕Diopatra,管口常用贝壳、海藻等碎屑伪装起来。②定居管栖类(sedentary tubicolou):口前叶具漏斗状的鳃冠或触手冠(brachial crown or tentacular c.)。鳃冠由许多鳃丝或触手丝(b.filament or radiole)组成,鳃丝上又常具两排鳃羽
枝(pinnule),躯干部分为胸腹两部分,见于龙介虫(石灰虫)(图18-1 K,图18-13
G)和缨鳃虫(图18-1 J)。③其他:磷虫Chaetopterus(图18-1 G,图18-15)具膜
状或牛皮纸状U形管,故又称parchment worm。笔帽虫(图18-1 H)栖管为上
细下粗,以细端外露于沙口的锥形管,由沙粒、海绵骨针、有孔虫壳或碎贝壳建
造而成。欧文虫栖管两端尖细,内壁角质富有弹性,粘有的沙粒或碎贝壳呈叠
瓦式排列。在帚毛虫,栖管常聚积在一起呈蜂窝状故又称honeycomb worm。
此外,阔沙蚕Platynereis等也具粘有沙粒或碎贝壳的临时性栖管。

图18-12 多毛动物(1)
A.吻沙蚕(吻翻出及其疣足);B.巧言虫(体前部吻翻出、体后部及疣足);
C.澳洲鳞沙蚕(体前部及外形);D.花索沙蚕(体前部及疣足);E.异足索沙蚕
(外形、体前部及疣足);F.矶沙蚕(体前部);G.扁犹帝虫(仙虫科)(外形、体前部、刚毛)

图18-13 多毛动物(2)
A.才女虫(海雅虫科)(虫体及体前部);B.须鳃虫(虫体及体前部);C.日本臭海蛹;
D.小头虫(虫体、雄性第8至第9节、巾钩刚毛); E.巴西沙蠋(虫体、刚毛);
F.不倒翁虫;G.龙介虫;H.吸口虫
(5)穿孔多毛类(boring polychaete):钻孔穴居于钙质贝壳、岩石或珊瑚中
的多毛类。如海稚虫科的才女虫Polydora常穿钻贝壳是珍珠贝养殖中的敌害
之一。丝鳃虫科的钙珊虫Dodecaceria(图18-16 B)常凿穴于钙质贝壳、珊瑚或
岩石中。
(6)小型多毛类(meiofaunal polychaete)(图18-14):分布于沉积物的沙间
间隙(沙间多毛类interstitial p.)或非沉积物的海藻、珊瑚、苔藓虫丛中或岩石
表面,为能通过1 mm(或0.5 mm)直径的筛网而保留于直径62μm(或41μm)
筛网上的多毛类(个别的小型多毛类如囊须虫却难以通过1 mm直径的筛网)。
小型多毛类常具细长的身体、细长的尾部和粘着器官(adhesive organ)或体节数
少、体常透明、常具纤毛头部附肢、疣足甚至刚毛退化不明显或缺少。习见的有
栉虫、沙虫、原须虫、好转虫、角虫、原虫。 此外沙蚕,裂虫等亦不乏有
小型者。因此,小型多毛类也不是个单一的分类类群。有人认为衍生自大型种
的退化或其幼态持续(paedomorphic)。

图18-14 小型多毛动物(仿Westheide等)
A.栉虫;B.沙虫;C.原须虫;D.好转虫;E.角虫(附尾部);F.囊须虫;G.原虫
(7)共栖(commensalism):多毛类的栖管、穴道里或鳞片下,常是其他动物
如小型蟹类、贝类甚至鳞沙蚕的寄宿场所。同时,有的多毛类又可生活于其他
动物如寄居蟹栖居的贝壳中或海星等的管足里。
(8)寄生(parasitism):寄生的多毛类不多见。寄花索沙蚕Labrorostratus parasiticus可寄生于其他多毛类的体腔中,最特殊的寄生者是吸口虫Myzostoma(图18-13 H)。
2.营养或摄食(nutrition or feeding):多毛类开拓了广泛的摄食小生境,其
摄食方式与栖息方式紧密相关。一部分向活动的肉食性或食腐性方向发展,另
一部分则向定居生活的等食或滤食方向演化。
(1)食肉者或捕食者(carnivore or raptorial feeder):包括爬行的、浮游的、
地下甬道穴居的和部分管栖者。沙蚕以翻吻和颚,矶沙蚕以复杂的颚器,裂虫
以齿或一圈小齿,吻沙蚕以翻吻和4个颚捕食和咀嚼,而有的游走管栖者甚至
能离开栖管外出。食物多为小型无脊椎动物亦包括其他多毛类。
(2)食草者、食腐肉者和啃食者(herbivore,scavenger and browser):具颚的
游走多毛类如沙蚕也兼食海藻,以其颚齿将海藻撕成碎片。有些矶沙蚕的颚器
更似刮刀。
(3)食泥沙者(sand and mud feeder):海滩中的细菌、硅藻、其他微小动物和
从水层中下落的有机碎屑,都是沉积物中有营养价值的食物资源。以此为生的
许多定居种类,可分为两类:①直接摄食沉积物者(direct deposite feeder),穴居
或管栖者,当他们通过穴道时,用来回外翻的囊状吻直接吞咽泥沙以获得食物。
其中有海蛹、小头虫、竹节虫和沙蠋等。锥头虫的树枝状吻可以舐食泥沙。②
间接摄食沉积物或触手摄食者(indirect deposite feeder or tentacle feeder),无
吻但具伸缩的触手,如大型穴居的蛰龙介和双栉虫的触手腹面具纤毛沟槽,可
将食物粒由纤毛沟送往触手基部聚积,而口侧的唇是食物进入口前的选择器
官。此外,海稚虫、丝鳃虫也都是靠有沟纤毛触手的间接摄食沉积物者。
(4)滤食或食悬浮物者(filter or suspension feeder):见于定居管栖类,能滤
食水层中悬浮的有机颗粒。头部具过滤食物粒的结构。常分为两类:①过渡型
滤食者(transitional filter feeder),有些海稚虫和欧文虫,不仅是底质沉积物的
间接摄食者而且也可摄食水中的悬浮颗粒。②真正的滤食者(true filter feeder),见于定居管栖类。帚毛虫壳盖柄腹侧长有摄食触手,被获食物沿一定路线
运送至口后被挑选。缨鳃虫和龙介虫的触手冠伸展时呈漏斗状,靠其纤毛的波
动把触手冠外的水流向中央集中,浮游生物或其他悬浮的有机颗粒便被滤出,
然后沿鳃丝的纤毛沟送往口中。在一种缨鳃虫Sabella davonina鳃丝的基部
形成深沟,起着挑选器(sorting device)的作用,只有最小的颗粒才落到沟底部
被送往口,中等颗粒在沟中被捕获运往腹囊用以建管或贮存,最大的颗粒保留
在沟的顶部,靠特殊的水流排出(图18-15 A,B)。
磷虫Chaetopterus(图18-15 C~F)是一种特殊的滤食动物——黏液过滤
摄食者(mucous feeding filter),其虫体结构高度变异,中区的圆扇叶(fans)像活
塞式地抽水入管,特别长的翼状叶(aliform)向背面弯曲成环,同时分泌黏液到
环中扩展成滤食的黏液袋(mucous bag),进入管中的水流和食物经黏液袋后被
俘获,于杯形器(cuporgan)中形成食物球,再通过背中部的纤毛沟往前行送至
口中,能滤食小到40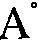 的颗粒,是过滤机制最有效的一种动物。 此外,磷虫也是
的颗粒,是过滤机制最有效的一种动物。 此外,磷虫也是
种发光生物,其生物(冷)光与裸甲藻、海萤、柱头虫等者不同,是发光蛋白在钙
离子作用下不需氧的发光体系:
![]()
3.生殖:多毛类生殖的多样性,在动物界也具代表性。
(1)无性生殖(asexual reproduction):断裂和出芽是多毛类中习见的无性
生殖。当与有性生殖有关时,断裂部分或出芽的芽体发育成有性个体。
1)断裂生殖(fragmentation):钙珊虫Dodecaceria(图18-16 B)、栉端虫
Ctenodrilus、叶磷虫Phyllochaetopterus、自裂虫Autolytus等,可自发地断裂成
单个体节或短的片断,断裂的部分可再生为一新个体,有的还可再重复产生一
系列的子个体,此为自发断裂(spontaneous f.)。另外,规则断裂(orderly f.)见
于尾稚虫Pygospio、斯氏缨鳃虫Sabella spallanzanii和裂虫科Syllidae、石灰
虫科Serpulidae的物种中,常在一定的部位横断(图18-17)。

图18-15 缨鳃虫和磷虫(仿各作者)
A~B.缨鳃虫:A.体前端示摄食流和建管,B.一个放射肋(鳃丝)的横切面示滤食;C~F.磷
虫:C.外形(背面观),D.在栖管中(示黏液袋的形成),E.再生(示由一个体节再生),F.发光

图18-16 星状钻穿裂虫的出芽出殖(A)和钙珊虫的再生(B)(仿各作者)

图18-17 多毛动物的无性生殖示意图(黑色示有性节,再生尾部以点表示)
A.齿裂虫Odontosyllis的异裂虫体,B.绿矶沙蚕Eunice viridis断裂的有性节区;
C.叉毛裂虫Syllis gracilis有性节放后,头部再生;D.粗毛裂虫S.amica有性节释
放前,头部再生;E.S.vitata有性节释放前,新头和无性节区新尾再生;F.释放第1个有
性节区前,第2个有性节区形成;G.自裂虫Autolytus多个匍枝的形成
(从各作者修改)
2)出芽生殖(budding,gemmiparity):由亲体的部分突起形成芽体的能力,
在多毛类中虽不多见,但变化却很大,尤其在裂虫科,如多育钻穿裂虫
Trypanosyllis prolifera几乎所有的体节都能产生头部,而且新个体的头链出
现于连续的体节上。星状钻穿裂虫T.asterobia(图18-16 A)可从单个体节上
出芽形成一丛有性个体。克氏钻穿裂虫T.crosslandi则只有肛节前产生一丛
有性个体。与深海六放海绵共栖的枝裂虫Syllis ramosa则在不同的体节上产
生侧生的不育芽体,芽体长大仍附于亲体的分枝群体上,仅在分开前,在有的侧
枝上产生次生的有性个体。
3)再生(regenaration):多毛类的触手、触角、触须、鳞片和其他柔弱的部分,
失去后会很快地再生出来。何况,体长的多毛类受捕食者的损害是常见现象。
磷虫Chaetopterus variopedatus(图18-15 C)和胶管虫Myxicola仅留1个体节
就能再生成完整的虫体。缨鳃虫Sabella的触手冠和胸部被鱼食去后,再生的
体节数最多不超过3节,其余的胸节则由腹部体节改造而成,此为变形再生
(morphallaxis)。尾部的再生率,常与被切去的体节数成正比。此外,磷虫极易
在翼状背叶和杯形器之间断开,显示其具保留重要的生殖区和当年生的体后
区,牺牲价值较小的体前部以供敌害果腹的自卫能力。
(2)有性生殖(sexual reproduction):
1)性(sex):多毛类多雌雄异体,但在裂虫、缨鳃虫、石灰虫等科的个别种中
具雌雄同体者。雌雄同体的缨鳃虫腹部前段产生卵而腹部后段产生精子。裂
虫有先雌后雄的,矶沙蚕科的眉轮虫Ophryotrocha有先雄后雌(雄性先熟protandrous)的性反转(sex reversa)现象。
2)生殖腺(gonad):多毛类生殖腺的中胚层来源是清楚的。但在不同科产
生于不同部位,除上述沙蚕科生殖腺来自腹隔膜体腔膜外,鳞虫科的鳞沙蚕
Aphrodita生殖腺则来自大血管处的体腔膜。原则上多数体节都可产生生殖细
胞,但明显分为胸腹部的多毛类,生殖腺常限于腹部。
3)生殖态(epitoky):有关生殖态的定义和形态变化在沙蚕中已做过介绍,
这常是沙蚕、裂虫和矶沙蚕为代表的特有的生殖现象,是由无性个体(非生殖个
体)向有性个体转变的过程,使之适于离开底穴、栖管或其他生境而起浮于水面
生殖(常在夜间、常具正的趋光性)。其形态变化包括头部、疣足和刚毛、体节的
大小以及体节肌等,所不同的是,有的直接转变(direct transformation)如沙蚕、
部分裂虫和矶沙蚕(图18-16 A)。有的匍枝繁殖(stolonization),即靠无性生殖
的断裂和出芽形成生殖匍枝(stolon),这常见于裂虫科(图18-18 B)。此外,多
型现象(polymorphism),常见于自裂虫Autolytus,其雌雄个体具不同的外形。

图18-18 沙蚕和裂虫生活史示意图
A.沙蚕;B.裂虫
最严格周期性群浮的例子,见于南太平洋萨摩亚群高潮下带、珊瑚或岩石
裂隙中生活的绿矶沙蚕Eunice viridis(Palolo viridis),其前部无性体区小,后
部有性体区由窄长且具腹眼的体节长链组成,在10月或11月的3/4月相的3
天里起浮且尤以第2天最甚。
多毛类的生殖靠激素调节。激素是由脑或前胃的神经成分(见于裂虫)产
生的神经分泌物。一生中只生殖一次的沙蚕或裂虫,激素既调节性细胞的产生
又调节生殖态性状发育的全部生殖过程。对多次生殖者,激素对性细胞特别是
对卵的发育是需要的,至于控制群浮等方面的准确机制仍知之甚少。
许多多毛类排卵于海水中,有些则将卵置于栖管、穴道或黏液团中,如巴西
沙蠋Arenicola brasiliensis等产卵袋于穴道口(图18-1 G)。有些种类,尤其是
定居管栖多毛类的卵多在管内孵化,螺旋虫Spirorbis可在壳盖腔中孵卵,自裂
虫Autolytus卵附于体腹面的囊孵化,极个别的种如盐沼沙蚕Nereis limnicola
的则在体腔中孵卵。
4)胚胎发育(embryogeny)和担轮幼虫(trochophora):多毛类的卵为多黄
卵、螺旋卵裂,虽在沙蚕、小头虫等类群发育为实囊胚(stereoblastula),但常具
可置换的囊胚腔。内陷、外包法完成原肠作用。原肠后胚胎很快发育为倒梨形
的担轮幼虫。 叶须虫、缨鳃虫、欧文虫和角虫的担轮幼虫为浮游营养(planktotrophic),而沙蚕、矶沙蚕者则卵黄营养(lecithotrophic),浮游期很短。
担轮幼虫(图18-19A,B)是多毛类等具螺旋卵裂原口动物的幼虫期,前端大于后端呈倒梨形。前端具1顶纤毛束(apical tuft),距顶端1/3~1/2处具围
绕身体的原纤毛轮(prototroch)或口前纤毛轮(preoral circle)。在多数多毛类
中第2个纤毛轮在口后发育,称后纤毛轮(metatroch)或口后纤轮(postoral c.),第3个纤毛轮位于后端肛门前称端纤毛轮(telotroch)。在内部,肠为完整
具纤毛的管,口位于原纤毛轮后,肠管和外胚层之间有的具囊胚腔且为幼虫肌
束穿过,一对原肾由两个中空的焰细胞组成、位于肠的两侧,排泄管开口于肛门
外侧,中胚层带位于腹侧、成体中胚层结构由此带发生。

图18-19 多毛纲的幼虫(仿各作者)
A.担轮幼虫外形;B.担轮幼虫结构;C.鳞沙蚕幼虫;D.欧文虫幼虫;E.磷虫幼虫;
F.石灰虫幼虫;G.海稚虫幼虫;H.担轮幼虫和成体各部间的关系
充分发育的担轮幼虫,可分为3区,并历经变态为成体:原毛轮区(prototrochal region)由顶纤毛束、原纤毛轮和口区组成,将来形成脑、口前叶和围口
节;肛区(anal r.)由端纤毛轮及其后的区域组成,系将来成体的肛节;生长带
(growth zone)位于口后纤毛轮后方和端纤毛轮前之间的区域,成体躯干部的全
部体节由此带逐渐分裂形成,使新体节在肛节前不断增多,因此最老的体节最
靠近头部。
担轮幼虫在动物界具很大的演化意义,他出现于涡虫纲的多肠目、软体动
物、环节动物多毛纲及其有关的类群。其早期阶段在许多方面是相似的,而明
显的差异只是在担轮幼虫后,这曾导致假想的担轮祖体理论(trochozoan theory)(Hatschek,1878),也对近代轮毛祖体理论(trochaea theory)的建立(Nielson等,1985)起了推动作用。