简介
简称DNA, 携带遗传信息的大分子。除少数RNA病毒的遗传物质是核糖核酸 (RNA) 外, DNA是所有其他生物的遗传物质。基因是DNA大分子上的一个区段(见基因)。
1953年, 美国J.瓦特森(Watson)和英国F. 克里克(Crick) 根据英国R. 富兰克林 (Franklin) 和H. 威尔金斯 (Wilkins) 对DNA分子的X射线衍射研究以及美国E. 贾格夫(Chargaff)对DNA碱基的化学分析提出了DNA的双螺旋结构模式。这个模式不仅从化学上肯定了基因的物质实体结构, 而且也为进一步了解基因的各种功能。如复制、突变、修复、重组、转录、翻译、调控以及人工合成、DNA体外重组等一系列重要分子生物学问题提供了依据, 从而为分子遗传学的诞生奠定了坚实的基础。
DNA是由两条互补的、极端细长的多聚脱氧核糖核酸链组成的。一条链的走向是5′→3′,另一条链的走向是3′→5′, 亦即这两条链是反向平行的, 彼此以一定的空向距离, 在同一轴上互相盘旋相交, 形成一个类似扭曲的梯子(见图1)。
每个脱氧核糖核酸又由脱氧核糖、磷酸和碱基组成。碱基有嘌呤和嘧啶之分, 前者为腺嘌呤(A)或鸟嘌呤(G), 后者为胞嘧啶(C)或胸腺嘧啶(T)。碱基A、G、C、T携带有遗传信息,因称之为遗传密码。而脱氧核糖和磷酸则主要起结构作用。脱氧核糖和磷酸位于分子外部, 它们以5′-3′磷酸二酯键相连, 成为分子的骨架, 而碱基则位于分子内部,以氢键相连。其中A永远与T配对, 中间有两个氢键; C永远与G配对, 中间有三个氢键,这就是碱基的互补配对规律。各碱基对之间的距离为3.4埃(Å),每段双螺旋内包含5个碱基对,双螺旋的直径为20埃。碱基间的氢键在温度接近100℃时或置入pH=3或pH=10的溶液中时, 可以断开, 使双链DNA分子分解成单链DNA, 称为变性。在温度逐渐变凉至65℃时,或溶液的pH恢复到中性时, 单链DNA根据碱基互补配对的规律, 可以重新形成双螺旋结构, 称为复性。四种碱基在双螺旋内不同的排列顺序就是大量不同排列组合的分子结构,亦即不同性质的基因。
绝大部分DNA分子的结构是右手螺旋, 通常以最常见的B- DNA为代表, 但在特殊条件下, 某些DNA的分子结构可以转变成左手螺旋,即Z-DNA。
DNA以其中一条链——有意义链为模板, 通过转录将其含有的遗传信息转变为mRNA, mRNA与另一条未转录DNA的无意义链的遗传信息一样, 只是其中以尿嘧啶(U)代替了T,于是mRNA上实际的遗传密码是A、U、C、G。接着, mRNA上的遗传密码经过翻译而形成蛋白质, 蛋白质就是生物性状的直接或间接体现者。翻译时每三个密码组成一个密码子, 翻译成一种氨基酸, 因称为三联体密码子。四个密码共有64种可能组合的密码子, 但UAA、UAG、UGA三个密码子为终止信号,并不翻译成氨基酸。其余61个密码子各翻译成一种氨基酸。但氨基酸只有20种, 因此一般是由两个以上的密码子共同翻译成一个氨基酸。此现象称为简并。各种氨基酸按遗传密码的顺序结合起来, 再经过折叠和卷曲就成为蛋白质。这就是DNA与蛋白质间的共线性关系。

图 1 DNA分子双螺旋结构模式
A. 腺嘌呤; T. 胸腺嘧啶; G. 鸟嘌呤; C. 胞嘧啶P. 磷酸; S. 脱氧核糖;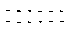 两个氢键;
两个氢键;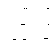 三个氢键;3.4Å两个碱基对之间的距离
三个氢键;3.4Å两个碱基对之间的距离
DNA作为遗传物质必然能进行自我复制, 这种复制首先是需要在双螺旋的一端打开氢键, 然后分别以每条老链作模板,各自形成与模板老链互补的新链。这样形成的两条链,各保留了一条原来的模板老链。因此称为半保留复制。
绝大多数生物DNA的复制是双向进行的, 只有极少数原核生物是单向复制的。原核生物的染色体有一个复制起点, 而真核生物每条染色体都有许多复制起点, 以保证必要的复制速度。
综合上述DNA的复制与蛋白质的合成过程 (即DNA的信息流向), 这就是分子生物学的中心法则(见图)。反转录酶和RNA自我复制的发现, 进一步丰富了这个法则。对深入理解生物遗传变异的实质有重要意义。
DNA分子的碱基顺序通常是相对固定的, 这体现了物种和品种的稳定性; 如果采用物理学或化学方法改变其碱基顺序或以碱基类似物代替某一碱基对,就会出现遗传性的变异——突变。

图2 中心法则示意
粗线示中心法则, 细线示最近发展,虚线示尚未发现的信息流向。
英文
deoxyribonucleic acid